欢迎访问作物学报,今天是
作物学报 ›› 2024, Vol. 50 ›› Issue (8): 2001-2013.doi: 10.3724/SP.J.1006.2024.33059
郭思语1,2( ), 赵克勇2(), 代正罡1,2, 邹华文1,*(), 吴忠义2,*(), 张春2,*()
), 赵克勇2(), 代正罡1,2, 邹华文1,*(), 吴忠义2,*(), 张春2,*()
GUO Si-Yu1,2(), ZHAO Ke-Yong2(), DAI Zheng-Gang1,2, ZOU Hua-Wen1,*(), WU Zhong-Yi2,*(), ZHANG Chun2,*()
摘要:
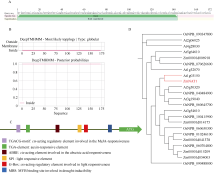
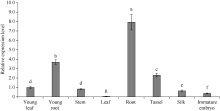
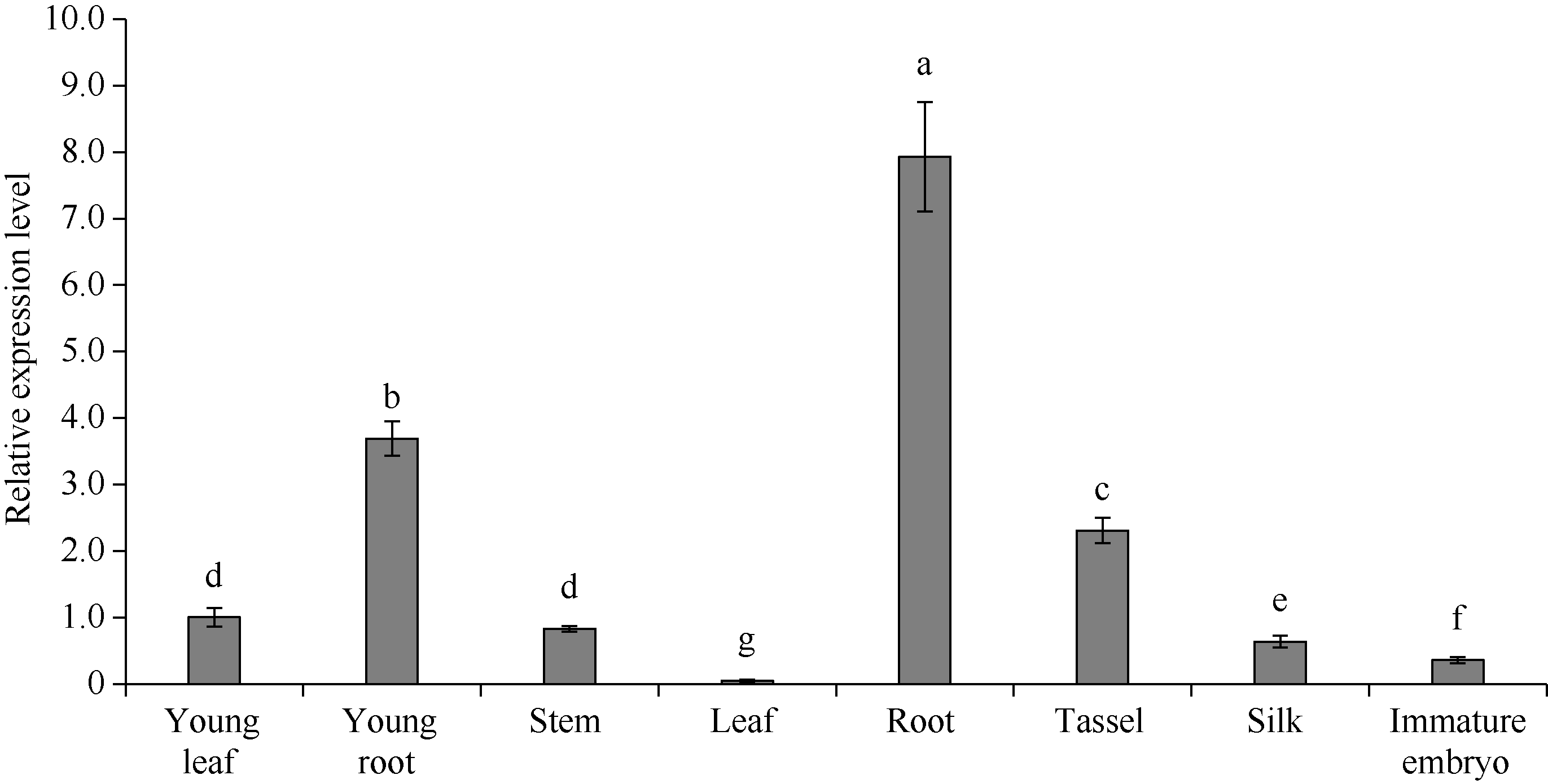
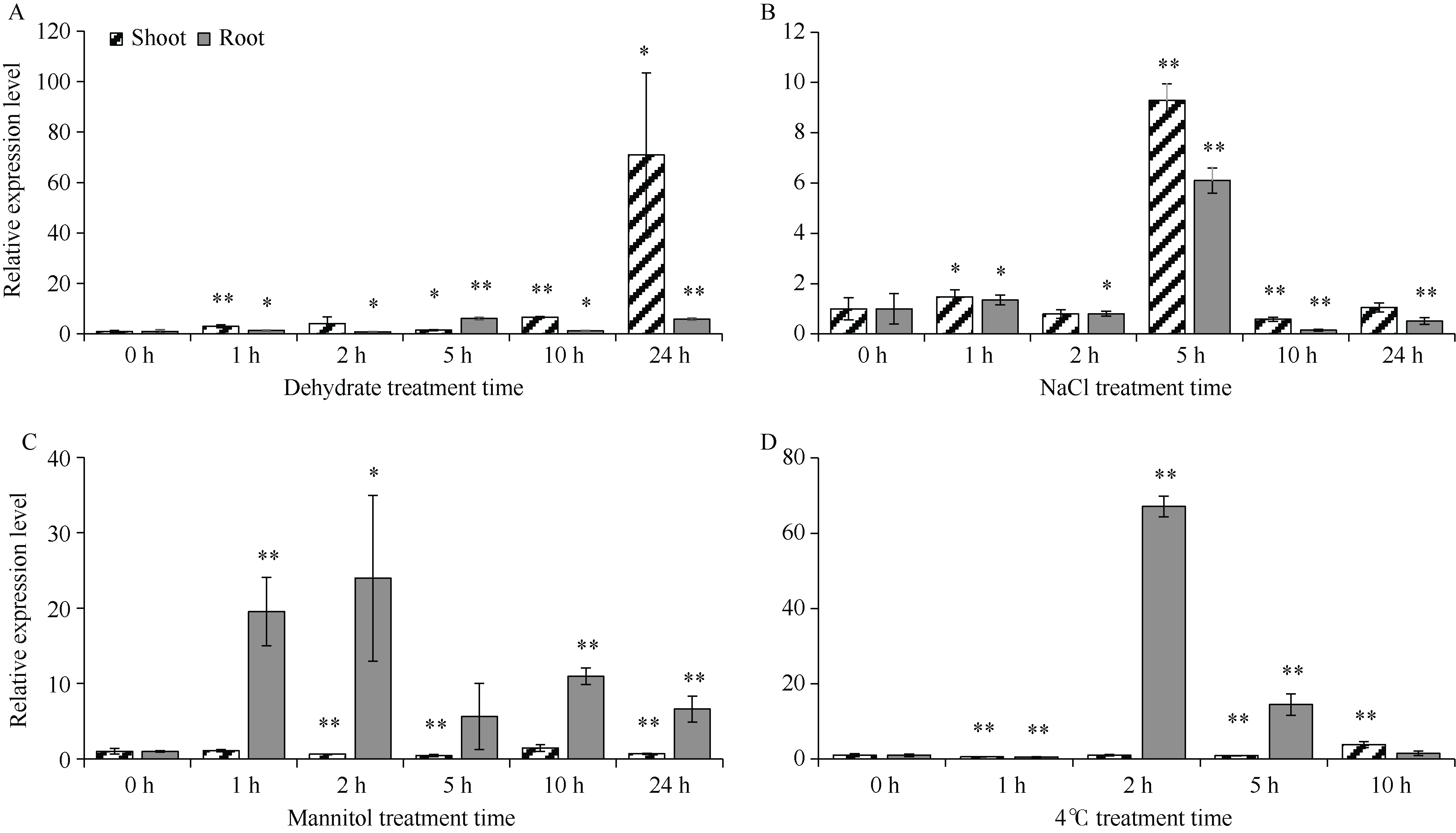
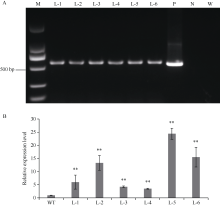
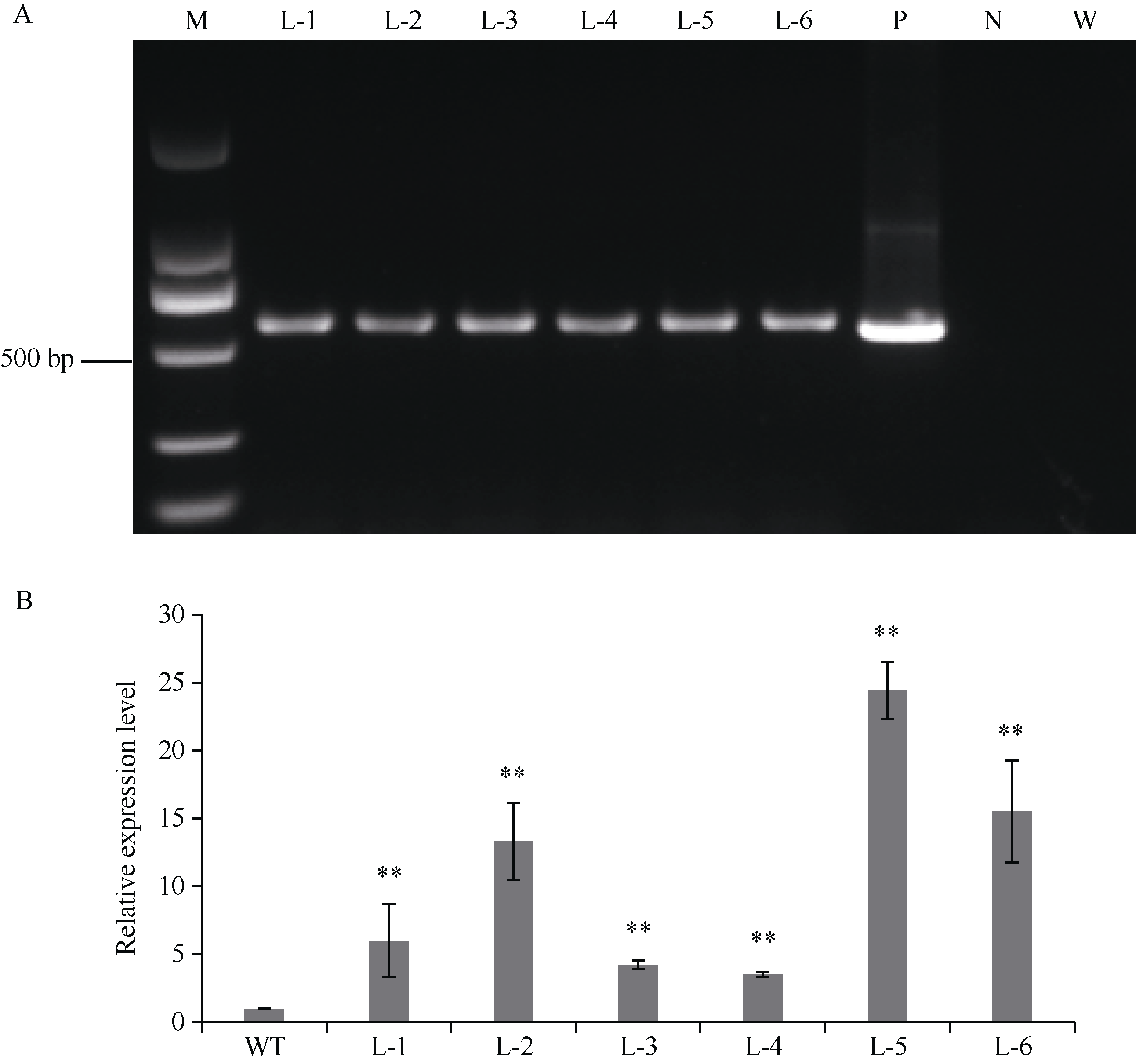
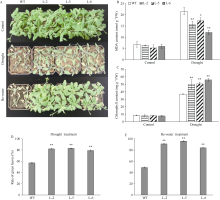
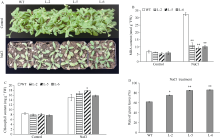
GNAT (Gcn5-related N-acetyltransferase)家族蛋白在调控植物生长发育和响应逆境胁迫等过程中发挥着重要作用。目前GNAT家族基因在多个物种中的生物学功能已有报道, 但在玉米(Zea mays L.)中的功能验证研究却很少。探究玉米GNAT家族基因的功能, 不仅能丰富我国的玉米育种基因资源, 同时也可为玉米的新种质资源创制提供重要依据。本研究克隆了ZmNAT1基因(Gene ID: 541936, GRMZM2G123159), 通过生物信息学分析发现, 该基因CDS全长为519 bp, 编码172个氨基酸, 具有GNAT家族特有的保守结构域。通过对ZmNAT1基因在玉米不同时期不同组织中的表达量和不同逆境胁迫下表达模式分析发现: ZmNAT1在成熟根中的表达量最高, 在不同非生物逆境胁迫处理下, ZmNAT1基因均有不同程度的诱导表达。通过异源表达获得了3株独立的表达量较高的转基因拟南芥(Arabidopsis thaliana L.)纯合株系, 对其进行了不同逆境胁迫处理下表型鉴定实验, 结果表明, 转基因拟南芥相对于野生型拟南芥有更好的表型, 在盐胁迫、渗透胁迫和干旱条件下的转基因株系的根显著长于野生型, 且植株较野生型植株的绿叶率和叶绿素含量均升高、丙二醛含量降低, 差异均达到显著水平。由此推测, ZmNAT1基因可能参与玉米对干旱、盐等非生物逆境胁迫的应答。本研究为进一步解析ZmNAT1在玉米中的生物学功能提供了重要的参考依据。
| [1] | Goharrizi K J, Meru G, Kermani S G, Heidarinezhad A, Salehi F. Short-term cold stress affects physiological and biochemical traits of pistachio rootstocks. S Afr J Bot, 2021, 141: 90-98. |
| [2] | 何含杰, 唐丽, 邓华凤, 邹志刚. 植物蛋白酪氨酸磷酸酶的生理功能研究进展. 植物生理学报, 2017, 53: 531-535. |
| He H J, Tang L, Deng H F, Zou Z G. Recent advance on the physiological functions of protein tyrosine phosphatases in plant. Acta Physiol Sin, 2017, 53: 531-535 (in Chinese with English abstract). | |
| [3] | 石永春, 杨永银, 薛瑞丽, 刘巧真. 植物中抗坏血酸的生物学功能研究进展. 植物生理学报, 2015, 51: 1-8. |
| Shi Y C, Yang Y Y, Xue R L, Liu Q Z. Research advance of biological functions of ascorbic acid in plants. Acta Physiol Sin, 2015, 51: 1-8 (in Chinese with English abstract). | |
| [4] | 王佺珍, 刘倩, 高娅妮, 柳旭. 植物对盐碱胁迫的响应机制研究进展. 生态学报, 2017, 37: 5565-5577. |
| Wang Q Z, Liu Q, Gao Y N, Liu X. Review on the mechanisms of response to salinity-alkalinity stress in plants. Acta Ecol Sin, 2017, 37: 5565-5577 (in Chinese with English abstract). | |
| [5] | 计淑霞, 戴绍军, 刘炜. 植物应答低温胁迫机制的研究进展. 生命科学, 2010, 22: 1013-1019. |
| Ji S X, Dai S J, Liu W. The advances of plants in response and adaption to low temperature stress. Life Sci, 2010, 22: 1013-1019 (in Chinese with English abstract). | |
| [6] | 冯鹏, 廖菲, 农向. 植物组蛋白乙酰转移酶研究进展. 乐山师范学院学报, 2022, 37(8): 8-14. |
| Feng P, Liao F, Nong X. Progress in plant histone acetyltransferases. J Leshan Norm Univ, 2022, 37(8): 8-14 (in Chinese). | |
| [7] |
Eberharter A, Becker P B. Histone acetylation: a switch between repressive and permissive chromatin. EMBO Rep, 2002, 3: 224-229.
doi: 10.1093/embo-reports/kvf053 pmid: 11882541 |
| [8] |
Kuo M H, Allis C D. Roles of histone acetyltransferases and deacetylases in gene regulation. Bioessays, 1998, 20: 615-626.
doi: 10.1002/(SICI)1521-1878(199808)20:8<615::AID-BIES4>3.0.CO;2-H pmid: 9780836 |
| [9] | Sterner D E, Berger S L. Acetylation of histones and transcription-related factors. Microbiol Mol Biol Rev, 2000, 64: 435-459. |
| [10] | Struhl K. Histone acetylation and transcriptional regulatory mechanisms. Genes Dev, 1998, 12: 599-606. |
| [11] |
Mai A, Rotili D, Tarantino D, Nebbioso A, Castellano S, Sbardella G, Tini M, Altucci L. Identification of 4-hydroxyquinolines inhibitors of p300/CBP histone acetyltransferases. Bioorg Med Chem Lett, 2009, 19: 1132-1135.
doi: 10.1016/j.bmcl.2008.12.097 pmid: 19144517 |
| [12] |
Kikuchi H, Nakayama T. GCN5 and BCR signalling collaborate to induce pre-mature B cell apoptosis through depletion of ICAD and IAP2 and activation of caspase activities. Gene, 2008, 419: 48-55.
doi: 10.1016/j.gene.2008.04.014 pmid: 18538956 |
| [13] |
Boycheva I, Vassileva V, Iantcheva A. Histone acetyltransferases in plant development and plasticity. Curr Genomics, 2014, 15: 28-37.
doi: 10.2174/138920291501140306112742 pmid: 24653661 |
| [14] |
Neuwald A F, Landsman D. GCN5-related histone N-acetyltransferases belong to a diverse superfamily that includes the yeast SPT10 protein. Trends Biochem Sci, 1997, 22: 154-155.
pmid: 9175471 |
| [15] |
夏德安, 刘春娟, 吕世博, 张彦妮, 刘奕佳, 马旭俊. 植物组蛋白乙酰基转移酶的研究进展. 生物技术通报, 2015, 31(7): 18-25.
doi: 10.13560/j.cnki.biotech.bull.1985.2015.07.003 |
|
Xia D A, Liu C J, Lyu S B, Zhang Y N, Liu Y J, Ma X J. Research progress of plant histone acetyltransferases. Biotechnol Bull, 2015, 31(7): 18-25 (in Chinese with English abstract).
doi: 10.13560/j.cnki.biotech.bull.1985.2015.07.003 |
|
| [16] | Servet C, Silva N C, Zhou D X. Histone acetyltransferase AtGCN5/HAG1 is a versatile regulator of developmental and inducible gene expression in Arabidopsis. Mol Plant, 2010, 3: 670-677. |
| [17] |
Bertrand C, Bergounioux C, Domenichini S, Delarue M, Zhou D X. Arabidopsis histone acetyltransferase AtGCN5 regulates the floral meristem activity through the WUSCHEL/AGAMOUS pathway. J Biol Chem, 2003, 278: 28246-28251.
doi: 10.1074/jbc.M302787200 pmid: 12740375 |
| [18] |
Cohen R, Schocken J, Kaldis A, Vlachonasios K E, Hark A T, Mccain E R. The histone acetyltransferase GCN5 affects the inflorescence meristem and stamen development in Arabidopsis. Planta, 2009, 230: 1207-1221.
doi: 10.1007/s00425-009-1012-5 pmid: 19771450 |
| [19] | Kornet N, Scheres B. Members of the GCN5 histone acetyltransferase complex regulate PLETHORA-mediated root stem cell niche maintenance and transit amplifying cell proliferation in Arabidopsis. Plant Cell, 2009, 21: 1070-1079. |
| [20] | Papaefthimiou D, Likotrafiti E, Kapazoglou A, Bladenopoulos K, Tsaftaris A. Epigenetic chromatin modifiers in barley: III. Isolation and characterization of the barley GNAT-MYST family of histone acetyltransferases and responses to exogenous ABA. Plant Physiol Bioch, 2010, 48: 98-107. |
| [21] | Fang H, Liu X, Thorn G, Duan J, Tian L. Expression analysis of histone acetyltransferases in rice under drought stress. Biochem Bioph Res Commun, 2014, 443: 400-405. |
| [22] |
Zheng M, Lin J C, Liu X B, Chu W, Li J P, Gao Y J, A K X, Song W J, Xin M M, Yao Y Y, Peng H R, Ni Z F, Sun Q X, Hu Z R. Histone acetyltransferase TaHAG1 acts as a crucial regulator to strengthen salt tolerance of hexaploid wheat. Plant Physiol, 2021, 186: 1951-1969.
doi: 10.1093/plphys/kiab187 pmid: 33890670 |
| [23] | Wang T Y, Xing J W, Liu Z S, Zheng M, Yao Y Y, Hu Z R, Peng H R, Xin M M, Zhou D X, Ni Z F. Histone acetyltransferase GCN5-mediated regulation of long non-coding RNA At4 contributes to phosphate starvation response in Arabidopsis. J Exp Bot, 2019, 70: 6337-6348. |
| [24] | Gan L, Wei Z Z, Yang Z R, Li F G, Wang Z. Updated mechanisms of GCN5: the monkey king of the plant kingdom in plant development and resistance to abiotic stresses. Cells, 2021, 10: 979. |
| [25] |
Eberharter A, Lechner T, Goralik-Schramel M, Loidl P. Purification and characterization of the cytoplasmic histone acetyltransferase B of maize embryos. FEBS Lett, 1996, 386: 75-81.
pmid: 8635608 |
| [26] | Lechner T, Lusser A, Brosch G, Eberharter A, Goralik-Schramel M, Loidl P. A comparative study of histone deacetylases of plant, fungal and vertebrate cells. Biochim Biophy Acta Prot Struct Mol Enzymolog, 1996, 1296: 181-188. |
| [27] |
Stockinger E J, Mao Y, Regier M K, Triezenberg S J, Thomashow M F. Transcriptional adaptor and histone acetyltransferase proteins in Arabidopsis and their interactions with CBF1, a transcriptional activator involved in cold-regulated gene expression. Nucleic Acids Res, 2001, 29: 1524-1533.
doi: 10.1093/nar/29.7.1524 pmid: 11266554 |
| [28] |
Bhat R A, Riehl M, Santandrea G, Velasco R, Slocombe S, Donn G, Steinbiss H H, Thompson R D, Becker H A. Alteration of GCN5 levels in maize reveals dynamic responses to manipulating histone acetylation. Plant J, 2003, 33: 455-469.
pmid: 12581304 |
| [29] | Georgieva E I, Lopez-Rodas G, Hittmair A, Feichtinger H, Brosch G, Loidl P. Maize embryo germination. Planta, 1993, 192: 118-124. |
| [30] |
Chang L, Loranger S S, Mizzen C, Ernst S G, Allis C D, Annunziato A T. Histones in transit: cytosolic histone complexes and diacetylation of H4 during nucleosome assembly in human cells. Biochemistry, 1997, 36: 469-480.
pmid: 9012662 |
| [31] |
马宇馨, 杜璇玥, 李肖慧, 任莹, 张林旺, 邢继红, 张康, 董金皋. 玉米组蛋白乙酰转移酶的鉴定与表达规律分析. 河北农业大学学报, 2020, 43(5): 20-26.
doi: 10.13320/j.cnki.jauh.2020.0089 |
| Ma Y X, Du X Y, Li X H, Ren Y, Zhang L W, Xing J H, Zhang K, Dong J G. Identification and expression analysis of histone acetyltransferase in maize. J Hebei Agric Univ, 2020, 43(5): 20-26 (in Chinese with English abstract). | |
| [32] | 余娇娇, 沈涛, 张晓东, 王雅妮. 缺氮胁迫下玉米组蛋白乙酰化相关酶的动态表达特征. 玉米科学, 2021, 29(6): 50-58. |
| Yu J J, Shen T, Zhang X D, Wang Y N. Dynamic expression patterns of corresponding enzymes of histone acetylation modification in maize under nitrogen deficiency. J Maize Sci, 2021, 29(6): 50-58 (in Chinese with English abstract). | |
| [33] |
悦曼芳, 张春, 郑登俞, 邹华文, 吴忠义. 玉米转录因子ZmbHLH91对非生物逆境胁迫的应答. 作物学报, 2022, 48: 3004-3017.
doi: 10.3724/SP.J.1006.2022.13060 |
| Yue M F, Zhang C, Zheng D Y, Zou H W, Wu Z Y. Response of maize transcriptional factor ZmbHLH91 to abiotic stress. Acta Agron Sin, 2022, 48: 3004-3017 (in Chinese with English abstract). | |
| [34] | Song Z T, Chen X J, Luo L, Yu F F, Liu J X, Han J J. UBA domain protein SUF1 interacts with NatA-complex subunit NAA15 to regulate thermotolerance in Arabidopsis. J Integr Plant Biol, 2022, 64: 1297-1302. |
| [35] |
Huber M, Bienvenut W V, Linster E, Stephan I, Armbruster L, Sticht C, Layer D, Lapouge K, Meinnel T, Sinning I, Giglione C, Hell R, Wirtz M. NatB-mediated N-terminal acetylation affects growth and biotic stress responses. Plant Physiol, 2020, 182: 792-806.
doi: 10.1104/pp.19.00792 pmid: 31744933 |
| [36] | Liu X, Luo M, Zhang W, Zhao J H, Zhang J X, Wu K Q, Tian L I, Duan J. Histone acetyltransferases in rice (Oryza sativa L.): phylogenetic analysis, subcellular localization and expression. BMC Plant Biol, 2012, 12: 145. |
| [37] | Fang H, Liu X, Thorn G, Duan J, Tian L. Expression analysis of histone acetyltransferases in rice under drought stress. Biochem Bioph Res Commun, 2014, 443: 400-405. |
| [38] |
Wang Z B, Zang C Z, Cui K R, Schones D E, Barski A, Peng W Q, Zhao K J. Genome-wide mapping of HATs and HDACs reveals distinct functions in active and inactive genes. Cell, 2009, 138: 1019-1031.
doi: 10.1016/j.cell.2009.06.049 pmid: 19698979 |
| [39] | Li S, Lin Y C J, Wang P Y, Zhang B F, Li M, Chen S, Shi R, Tunlaya-Anukit S, Liu X Y, Wang Z F, Dai X F, Yu J, Zhou C G, Liu B G, Wang J P, Chiang V L, Li W. The AREB1 transcription factor influences histone acetylation to regulate drought responses and tolerance in Populus trichocarpa. Plant Cell, 2019, 31: 663-686. |
| [40] |
Bertrand C, Benhamed M, Li Y F, Ayadi M, Lemonnier G, Renou J P, Delarue M, Zhou D X. Arabidopsis HAF2 gene encoding TATA-binding protein (TBP)-associated factor TAF1, is required to integrate light signals to regulate gene expression and growth. J Biol Chem, 2005, 280: 1465-1473.
doi: 10.1074/jbc.M409000200 pmid: 15525647 |
| [41] |
Li H, Yan S H, Zhao L, Tan J J, Zhang Q, Gao F, Wang P, Hou H L, Li L J. Histone acetylation associated up-regulation of the cell wall related genes is involved in salt stress induced maize root swelling. BMC Plant Biol, 2014, 14: 105.
doi: 10.1186/1471-2229-14-105 pmid: 24758373 |
| [1] | 叶靓, 朱叶琳, 裴琳婧, 张思颖, 左雪倩, 李正真, 刘芳, 谭静. 联合全基因组关联和转录组分析筛选玉米拟轮枝镰孢穗腐病的抗性候选基因[J]. 作物学报, 2024, 50(9): 2279-2296. |
| [2] | 杨煜琛, 靳雅荣, 骆金婵, 祝鑫, 李葳航, 贾纪原, 王小珊, 黄德均, 黄琳凯. 珍珠粟WD40基因家族鉴定及表达特征分析[J]. 作物学报, 2024, 50(9): 2219-2236. |
| [3] | 孙照华, 任昊, 王洪章, 王子强, 姚海燕, 辛爱美, 赵斌, 张吉旺, 任佰朝, 刘鹏. 叶面喷施硅制剂对滨海盐碱地夏玉米叶片光合性能及籽粒产量的影响[J]. 作物学报, 2024, 50(9): 2383-2395. |
| [4] | 曹晓晴, 祁显涛, 刘昌林, 谢传晓. 编辑ZmCCT10、ZmCCT9、ZmGhd7基因的串联DsRed荧光表达盒的CRISPR/Cas9系统的构建及验证[J]. 作物学报, 2024, 50(8): 1961-1970. |
| [5] | 刘陈, 王昆昆, 廖世鹏, 杨佳群, 丛日环, 任涛, 李小坤, 鲁剑巍. 氮肥用量对玉米-油菜和水稻-油菜轮作模式下油菜产量及氮素吸收利用的影响[J]. 作物学报, 2024, 50(8): 2067-2077. |
| [6] | 刘宸铭, 赵克勇, 悦曼芳, 赵延明, 吴忠义, 张春. 玉米转录因子ZmEREB180调控根系生长发育及耐逆的功能研究[J]. 作物学报, 2024, 50(8): 1920-1933. |
| [7] | 刘爽, 李珅, 王东梅, 沙小茜, 何冠华, 张登峰, 李永祥, 刘旭洋, 王天宇, 黎裕, 李春辉. 基于大刍草渗入系的玉米抗旱优异等位基因挖掘[J]. 作物学报, 2024, 50(8): 1896-1906. |
| [8] | 梁璐, 周宝元, 高卓晗, 王瑞, 王新兵, 赵明, 李从锋. 不同品种玉米根-冠生长对土壤紧实胁迫的差异性响应特征[J]. 作物学报, 2024, 50(8): 2053-2066. |
| [9] | 方宇辉, 齐学礼, 李艳, 张煜, 彭超军, 华夏, 陈艳艳, 郭瑞, 胡琳, 许为钢. 强光胁迫对转玉米C4型ZmPEPC+ZmPPDK基因小麦光合和生理特性的影响[J]. 作物学报, 2024, 50(7): 1647-1657. |
| [10] | 韩笑晨, 张贵芹, 王亚辉, 任昊, 王洪章, 刘国利, 林佃旭, 王子强, 张吉旺, 赵斌, 任佰朝, 刘鹏. 土壤调理剂对滨海盐碱地土壤盐分含量及夏玉米产量的影响[J]. 作物学报, 2024, 50(7): 1776-1786. |
| [11] | 王蕊, 孙擘, 张云龙, 张茗起, 范亚明, 田红丽, 赵怡锟, 易红梅, 匡猛, 王凤格. 叶绿体标记在玉米种质资源快速分组中的应用分析[J]. 作物学报, 2024, 50(7): 1867-1876. |
| [12] | 唐清芸, 杨晶晶, 赵蕾, 宋志文, 王国栋, 李玉祥. 施氮量对滴灌水稻根系形态构型和分形特征的影响[J]. 作物学报, 2024, 50(6): 1540-1553. |
| [13] | 王菲儿, 郭瑶, 李盼, 韦金贵, 樊志龙, 胡发龙, 范虹, 何蔚, 殷文, 陈桂平. 绿洲灌区增密对水氮减量玉米产量的补偿机制[J]. 作物学报, 2024, 50(6): 1616-1627. |
| [14] | 折萌, 郑登俞, 柯照, 吴忠义, 邹华文, 张中保. 玉米ZmGRAS13基因的克隆及功能研究[J]. 作物学报, 2024, 50(6): 1420-1434. |
| [15] | 郑雪晴, 王兴荣, 张彦军, 龚佃明, 邱法展. 玉米果穗相关性状QTL定位及重要候选基因分析[J]. 作物学报, 2024, 50(6): 1435-1450. |
|