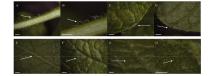
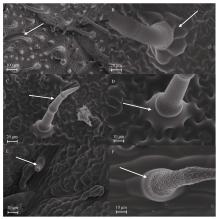
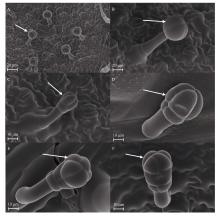
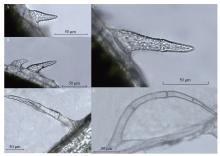
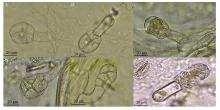
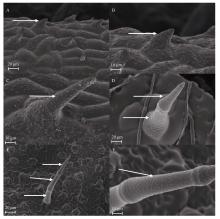
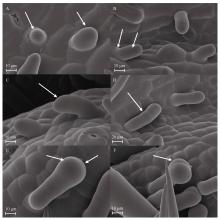
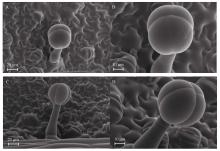
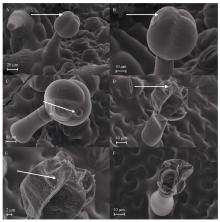
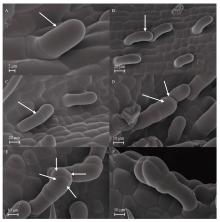
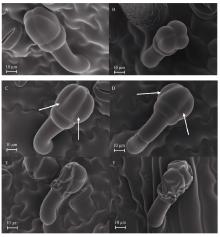
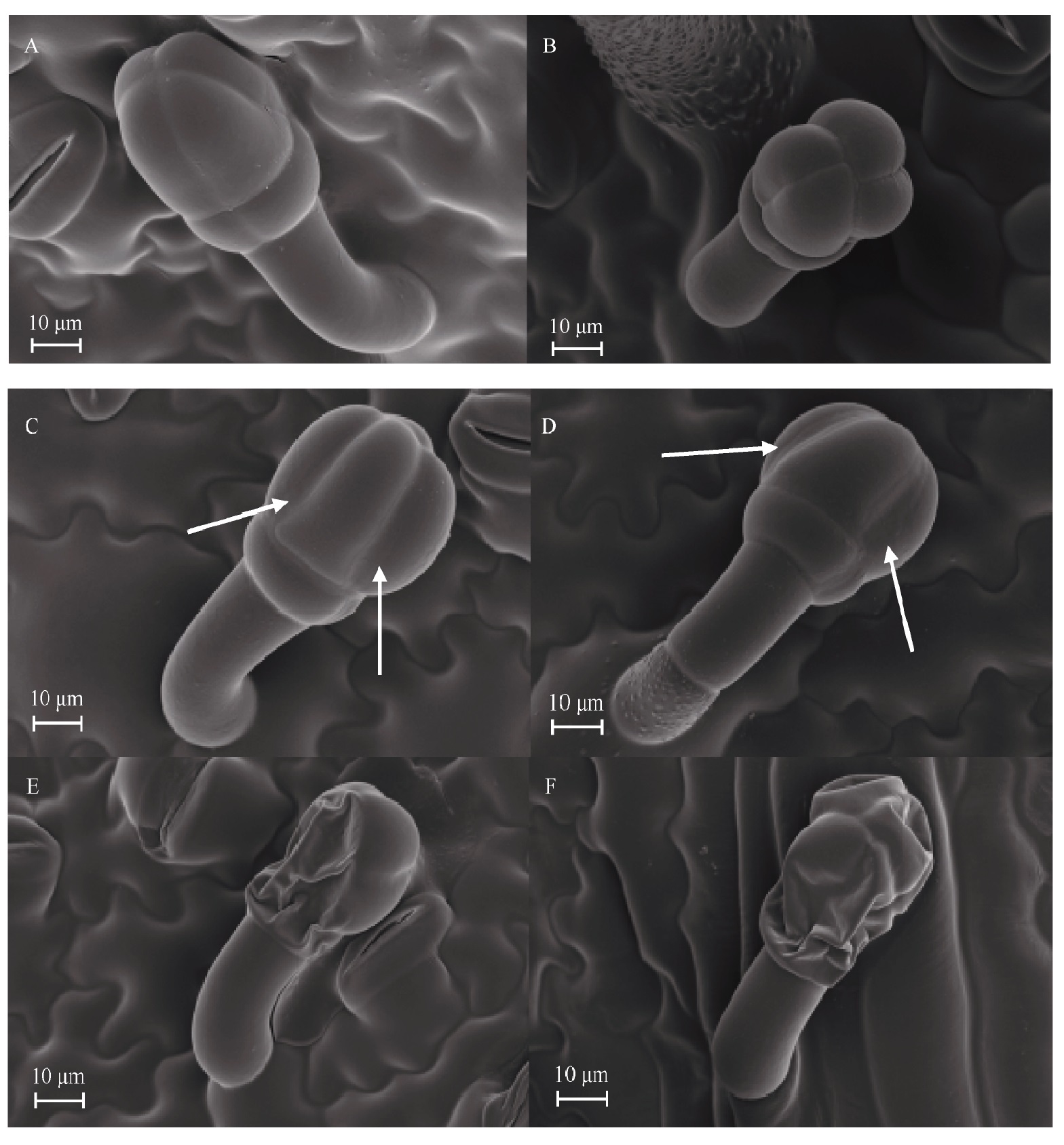
Welcome to Acta Agronomica Sinica,
Acta Agronomica Sinica ›› 2025, Vol. 51 ›› Issue (6): 1582-1598.doi: 10.3724/SP.J.1006.2025.44188
• TILLAGE & CULTIVATION·PHYSIOLOGY & BIOCHEMISTRY • Previous Articles Next Articles
YANG Shuang( ), BAI Lei, GUO Hua-Chun, MIAO Ya-Sheng, LI Jun()
), BAI Lei, GUO Hua-Chun, MIAO Ya-Sheng, LI Jun()
| [1] | 马璐璐, 刘昭, 陈佳琪, 李伟, 金晓霞, 岳中辉. 龙葵腺毛发育的细胞学研究. 西北植物学报, 2023, 43: 1959-1968. |
| Ma L L, Liu Z, Chen J Q, Li W, Jin X X, Yue Z H. Studies on cytology of glandular trichomes of Solanum nigrum L. Acta Bot Boreali-Occident Sin, 2023, 43: 1959-1968 (in Chinese with English abstract). | |
| [2] |
Kaur S, Khanal N, Dearth R, Kariyat R. Morphological characterization of intraspecific variation for trichome traits in tomato (Solanum lycopersicum). Bot Stud, 2023, 64: 7.
doi: 10.1186/s40529-023-00370-3 pmid: 36988701 |
| [3] | Lustofin K, Świątek P, Miranda V F O, Płachno B J. Flower nectar trichome structure of carnivorous plants from the genus butterworts Pinguicula L. (Lentibulariaceae). Protoplasma, 2020, 257: 245-259. |
| [4] | Qin W, Li Y P, Peng B W, Liu H, Chen T T, Yan X, Zhang Y J, Wang C, Yao X H, Fu X Q, et al. A high-efficiency trichome collection system by laser capture microdissection. Front Plant Sci, 2022, 13: 985969. |
| [5] | Chen C H, Liu M L, Jiang L, Liu X F, Zhao J Y, Yan S S, Yang S, Ren H Z, Liu R Y, Zhang X L. Transcriptome profiling reveals roles of meristem regulators and polarity genes during fruit trichome development in cucumber (Cucumis sativus L.). J Exp Bot, 2014, 65: 4943-4958. |
| [6] | 左美娜.蔷薇属植物腺体型皮刺的形态结构及发育规律. 华中农业大学硕士学位论文, 湖北武汉, 2023. |
| Zuo M N. Morphological Structure and Developmental Regularity of Glandular Prickles of the Rosa genus. MS Thesis of Huazhong Agricultural University, Wuhan, Hubei, China, 2023 (in Chinese with English abstract). | |
| [7] | 杨宗燃.二色补血草MYB转录因子LbMYB17的功能研究. 山东师范大学硕士学位论文, 山东济南, 2024. |
| Yang Z R. Functional Study on MYB Transcription Factor LbMYB17 of Limonium bicolor. MS Thesis of Shandong Normal University, Jinan, Shandong, China, 2024 (in Chinese with English abstract). | |
| [8] | 王露, 赵天祎, 蔡明. 植物腺毛中防御物质的合成与调控及转运研究进展. 西北植物学报, 2021, 41: 338-347. |
| Wang L, Zhao T Y, Cai M. Advances in synthesis, regulation and transportation of defensive substances produced in plant glandular trichomes. Acta Bot Boreali-Occident Sin, 2021, 41: 338-347 (in Chinese with English abstract). | |
| [9] |
Glas J J, Schimmel B C J, Alba J M, Escobar-Bravo R, Schuurink R C, Kant M R. Plant glandular trichomes as targets for breeding or engineering of resistance to herbivores. Int J Mol Sci, 2012, 13: 17077-17103.
doi: 10.3390/ijms131217077 pmid: 23235331 |
| [10] | Amrehn E, Spring O. Ultrastructural alterations in cells of sunflower linear glandular trichomes during maturation. Plants, 2021, 10: 1515. |
| [11] | Qi X W, Chen Z Q, Yu X, Li L, Bai Y, Fang H L, Liang C Y. Characterisation of the Mentha canadensis R2R3-MYB transcription factor gene McMIXTA and its involvement in peltate glandular trichome development. BMC Plant Biol, 2022, 22: 219. |
| [12] | 林宗梅.NtCAF1影响拟南芥表皮毛发育的功能研究. 贵州大学硕士毕业论文, 贵州贵阳, 2021. |
| Lin Z M. Functional Study of NtCAF1on the Development of Trichomes in Arabidopsis. MS Thesis of Guizhou University, Guiyang, Guizhou, China, 2021 (in Chinese with English abstract). | |
| [13] | 曹可心, 杨惠娟, 刘化冰, 吴健, 陈小翔, 徐世晓. NtCPS2基因通过调控赤霉素合成影响烟草腺毛发生. 农业生物技术学报, 2024, 32: 1784-1791. |
| Cao K X, Yang H J, Liu H B, Wu J, Chen X X, Xu S X. NtCPS2 gene affects the occurrence of tobacco (Nicotiana tabacum) glandular hair by regulating gibberellin synthesis. J Agric Biotechnol, 2024, 32: 1784-1791 (in Chinese with English abstract). | |
| [14] | Watts S, Kariyat R. Morphological characterization of trichomes shows enormous variation in shape, density and dimensions across the leaves of 14 Solanum species. AoB Plants, 2021, 13: plab071. |
| [15] | 李洁, 张铭芳, 于见丽, 张秀海, 杜运鹏, 王庆江. 植物表皮毛发育调控的研究进展. 北方园艺, 2020, (23): 133-139. |
| Li J, Zhang M F, Yu J L, Zhang X H, Du Y P, Wang Q J. Advances in regulation of plant trichome development. North Hortic, 2020, (23): 133-139 (in Chinese with English abstract). | |
| [16] | 钟姗辰, 武舒, 王黎, 苏晓华, 张冰玉. 植物毛状体研究进展. 分子植物育种, 2023, 21: 7215-7224. |
| Zhong S C, Wu S, Wang L, Su X H, Zhang B Y.Advances in molecular mechanisms of plant Trichomonas. Mol Plant Breed, 2023, 21: 7215-7224 (in Chinese with English abstract). | |
| [17] | 方小婷, 李佩华, 兰建彬, 赵飞, 王芳, 郑顺林.植物毛状体功能与马铃薯毛状体研究.见: 金黎平, 吕文河主编. 马铃薯产业与大食物观(2024). 中国作物学会马铃薯专业委员会, 2024. pp 142-150. |
| Fang X T, Li P H, Lang J B, Zhao F, Wang F, Zheng S L. Plant trichome function and potato trichome research, In: Jin L P, Lu W H, eds. Potato Industry and Big Food Concept (2024). Potato Committee of Crop Science Society of China, 2024. pp 142-150 (in Chinese). | |
| [18] | Papierowska E, Szatyłowicz J, Samborski S, Szewińska J, Różańska E. The leaf wettability of various potato cultivars. Plants, 2020, 9: 504. |
| [19] | Yan A, Pan J B, An L Z, Gan Y B, Feng H Y.The responses of trichome mutants to enhanced ultraviolet-B radiation in Arabidopsis thaliana. J Photochem Photobiol B, 2012, 113: 29-35. |
| [20] | Shukla P, Kidwai M, Narayan S, Shirke P A, Pandey K D, Misra P, Chakrabarty D. Phytoremediation potential of Solanum viarum Dunal and functional aspects of their capitate glandular trichomes in lead, cadmium, and zinc detoxification. Environ Sci Pollut Res Int, 2023, 30: 41878-41899. |
| [21] |
Liu Y, Jing S X, Luo S H, Li S H. Non-volatile natural products in plant glandular trichomes: chemistry, biological activities and biosynthesis. Nat Prod Rep, 2019, 36: 626-665.
doi: 10.1039/c8np00077h pmid: 30468448 |
| [22] | 毛静, 杨品, 周媛, 童俊, 徐冬云. 杜鹃叶片表皮毛特征对杜鹃冠网蝽寄主选择的影响. 江苏农业科学, 2022, 50(20): 154-160. |
| Mao J, Yang P, Zhou Y, Tong J, Xu D Y. Influence of rhododendron leaf trichomes characteristics on host selection of Stephanitis pyriodes Scott. Jiangsu Agric Sci, 2022, 50(20): 154-160 (in Chinese). | |
| [23] | Munien P, Naidoo Y, Naidoo G. Micromorphology, histochemistry and ultrastructure of the foliar trichomes of Withania somnifera (L.) Dunal (Solanaceae). Planta, 2015, 242: 1107-1122. |
| [24] |
Rodziewicz P, Loroch S, Marczak Ł, Sickmann A, Kayser O. Cannabinoid synthases and osmoprotective metabolites accumulate in the exudates of Cannabis sativa L. glandular trichomes. Plant Sci, 2019, 284: 108-116.
doi: S0168-9452(18)31486-9 pmid: 31084863 |
| [25] | 亓超凡, 刘艳华, 刘静, 杜咏梅, 刘新民, 韩晓, 雷云康, 张洪博, 付秋娟, 欧阳一鸣, 等. 植物腺毛分泌物研究进展. 福建农业学报, 2024, 39(1): 115-124. |
| Qi C F, Liu Y H, Liu J, Du Y M, Liu X M, Han X, Lei Y K, Zhang H B, Fu Q J, Ou-Yang Y M, et al. Research progress on glandular trichome secretions of plants. Fujian J Agric Sci, 2024, 39(1): 115-124 (in Chinese with English abstract). | |
| [26] | 邸慧慧, 史宏志, 张大纯. 烟草叶片腺毛及其分泌物研究进展. 贵州农业科学, 2009, 37(8): 61-64. |
| Di H H, Shi H Z, Zhang D C. Research progress on glandular hair and glandular hair secretion of tobacco leaves. Guizhou Agric Sci, 2009, 37(8): 61-64 (in Chinese with English abstract). | |
| [27] | 刘孟奇, 回成程, 陈随清, 郭涛, 王海波, 辛海量. 艾叶及其腺毛显微鉴定研究. 中草药, 2022, 53: 7532-7542. |
| Liu M Q, Hui C C, Chen S Q, Guo T, Wang H B, Xin H L. Microscopic identification and histochemical study of glandular trichomes in Artemisia argyi leaves. Chin Tradit Herb Drugs, 2022, 53: 7532-7542 (in Chinese with English abstract). | |
| [28] |
宋倩娜, 宋慧洋, 李京昊, 段永红, 梅超, 冯瑞云. 马铃薯转录因子StFBH3对非生物逆境胁迫的响应分析. 作物学报, 2025, 51: 247-259.
doi: 10.3724/SP.J.1006.2025.44044 |
| Song Q N, Song H Y, Li J H, Duan Y M, Mei C, Feng R Y. Response of transcription factor StFBH3 under abiotic stress in potato. Acta Agron Sin, 2025, 51: 247-259 (in Chinese with English abstract). | |
| [29] |
Qu L, Huang X Q, Su X, Zhu G Q, Zheng L L, Lin J, Wang J W, Xue H W. Potato: from functional genomics to genetic improvement. Mol Hortic, 2024, 4: 34.
doi: 10.1186/s43897-024-00105-3 pmid: 39160633 |
| [30] | 刘天天, 杨迪, 孙廷, 惠丰立, 常换换, 宋玉伟. 植物非腺毛形态、功能及应用研究进展. 分子植物育种, 网络首发 [2023-03-01], https://kns.cnki.net/kcms/detail//46.1068.S.20230301.1045.009.html. . |
| Liu T T, Yang D, Sun T, Hui F L, Chang H H, Song Y W. Research progress on morphology, function and application of plant non-glandular trichomes. Mol Plant Breed, Published online , [2023-03-01], https://kns.cnki.net/kcms/detail//46.1068.S.20230301.1045.009.html. (in Chinese with English abstract). | |
| [31] | Li C, Wu J T, Blamey F P C, Wang L L, Zhou L N, Paterson D J, van der Ent A, Fernández V, Lombi E, Wang Y H, et al. Non-glandular trichomes of sunflower are important in the absorption and translocation of foliar-applied Zn. J Exp Bot, 2021, 72: 5079-5092. |
| [32] | Kang J H, Shi F, Daniel Jones A, David Marks M, Howe G A. Distortion of trichome morphology by the hairless mutation of tomato affects leaf surface chemistry. J Exp Bot, 2010, 61: 1053-1064. |
| [33] |
McDowell E T, Kapteyn J, Schmidt A, Li C, Kang J H, Descour A, Shi F, Larson M, Schilmiller A, An L L, et al. Comparative functional genomic analysis of Solanum glandular trichome types. Plant Physiol, 2011, 155: 524-539.
doi: 10.1104/pp.110.167114 pmid: 21098679 |
| [34] |
Wang F M, Park Y L, Gutensohn M. Glandular trichome-derived sesquiterpenes of wild tomato accessions (Solanum habrochaites) affect aphid performance and feeding behavior. Phytochemistry, 2020, 180: 112532.
doi: 10.1016/j.phytochem.2020.112532 pmid: 33045464 |
| [35] |
Fan P X, Leong B J, Last R L. Tip of the trichome: evolution of acylsugar metabolic diversity in Solanaceae. Curr Opin Plant Biol, 2019, 49: 8-16.
doi: S1369-5266(18)30090-6 pmid: 31009840 |
| [36] |
Luu V T, Weinhold A, Ullah C, Dressel S, Schoettner M, Gase K, Gaquerel E, Xu S Q, Baldwin I T. O-acyl sugars protect a wild tobacco from both native fungal pathogens and a specialist herbivore. Plant Physiol, 2017, 174: 370-386.
doi: 10.1104/pp.16.01904 pmid: 28275149 |
| [37] | Rodriguez A M, Derita M G, Andrada A R, de los A Páez V, Ponce M, Martínez O G, Neira D A, Hernández M A. Glandular trichomes as a source of antifungal metabolites in a karyologically characterized population of Argyrochosma flava: morphology and histochemistry. Flora, 2024, 311: 152456. |
| [38] | Arafa R A, Kamel S M, Taher D I, Solberg S Ø, Rakha M T. Leaf extracts from resistant wild tomato can be used to control late blight (Phytophthora infestans) in the cultivated tomato. Plants, 2022, 11: 1824. |
| [39] | Gorzolka K, Perino E H B, Lederer S, Smolka U, Rosahl S. Lysophosphatidylcholine 17: 1 from the leaf surface of the wild potato species Solanum bulbocastanum inhibits Phytophthora infestans. J Agric Food Chem, 2021, 69: 5607-5617. |
| [40] |
Szymanski D B, Jilk R A, Pollock S M, Marks M D. Control of GL2 expression in Arabidopsis leaves and trichomes. Development, 1998, 125: 1161-1171.
doi: 10.1242/dev.125.7.1161 pmid: 9477315 |
| [41] |
Nanayakkara U N, Nie X, Giguère M, Zhang J, Boquel S, Pelletier Y. Aphid feeding behavior in relation to potato virus Y (PVY) acquisition. J Econ Entomol, 2012, 105: 1903-1908.
pmid: 23356052 |
| [42] | Murphy A F, Rondon S I, Moreno A, Fereres A. Effect of potato virus Y presence in Solanum tuberosum (Solanales: Solanaceae) and Chenopodium album on aphid (Hemiptera: Aphididae) behavior. Environ Entomol, 2018, 47: 654-659. |
| [43] | Horgan F G, Quiring D T, Lagnaoui A, Pelletier Y. Effects of altitude of origin on trichome-mediated anti-herbivore resistance in wild Andean potatoes. Flora Morphol Distrib Funct Ecol Plants, 2009, 204: 49-62. |
| [44] | 马中正, 任彬元, 赵中华, 李春广, 沈杰, 闫硕. 近年我国马铃薯四大产区病虫害发生及防控情况的比较分析. 植物保护学报, 2020, 47: 463-470. |
| Ma Z Z, Ren B Y, Zhao Z H, Li C G, Shen J, Yan S. Comparative analysis of the occurrence and control of pests and diseases in four major potato producing areas in China in recent years. J Plant Prot, 2020, 47: 463-470 (in Chinese with English abstract). | |
| [45] |
徐进, 朱杰华, 杨艳丽, 汤浩, 吕和平, 樊明寿, 石瑛, 董道峰, 王贵江, 王万兴, 等. 中国马铃薯病虫害发生情况与农药使用现状. 中国农业科学, 2019, 52: 2800-2808.
doi: 10.3864/j.issn.0578-1752.2019.16.006 |
|
Xu J, Zhu J H, Yang Y L, Tang H, Lyu H P, Fan M S, Shi Y, Dong D F, Wang G J, Wang W X, et al. Status of major diseases and insect pests of potato and pesticide usage in China. Sci Agric Sin, 2019, 52: 2800-2808 (in Chinese with English abstract).
doi: 10.3864/j.issn.0578-1752.2019.16.006 |
|
| [46] |
Crossley M S, Schoville S D, Haagenson D M, Jansky S H. Plant resistance to Colorado potato beetle (Coleoptera: Chrysomelidae) in diploid F2 families derived from crosses between cultivated and wild potato. J Econ Entomol, 2018, 111: 1875-1884.
doi: 10.1093/jee/toy120 pmid: 29688507 |
| [47] | Molnar I, Rakosy-Tican E. Difficulties in potato pest control: the case of pyrethroids on Colorado potato beetle. Agronomy, 2021, 11: 1920. |
| [1] | . Effects of ionic zinc and nano-zinc on physiological characteristics, yield, and quality of potato [J]. Acta Agronomica Sinica, 2025, 51(7): 1838-1849. |
| [2] | YIN Yu-Meng, WANG Yan-Nan, KANG Zhi-He, QIAO Shou-Chen, BIAN Qian-Qian, LI Ya-Wei, CAO Guo-Zheng, ZHAO Guo-Rui, XU Dan-Dan, YANG Yu-Feng. Cloning and functional analysis of glutathione S-transferase gene IbGSTU7 in sweetpotato [J]. Acta Agronomica Sinica, 2025, 51(7): 1736-1746. |
| [3] | SHAO Shun-Wei, CHEN Zhuo, LAN Zhen-Dong, CAI Xing-Kui, ZOU Hua-Fen, LI Chen-Xi, TANG Jing-Hua, ZHU Xi, ZHANG Yu, DONG Jian-Ke, JIN Hui, SONG Bo-Tao. QTL mapping of tuber eye depth based on BSA-seq technique [J]. Acta Agronomica Sinica, 2025, 51(7): 1725-1735. |
| [4] | YUAN Xin, ZHAO Zhuo-Fan, ZHAO Rui-Qing, LIU Xiao-Wei, ZHENG Ming-Min, LIU Yu-Sheng, DONG Hao-Sheng, DENG Li-Juan, CAO Mo-Ju, HUANG Qiang. Genetic analysis and molecular identification of a small kernel mutant mn-like1 in maize [J]. Acta Agronomica Sinica, 2025, 51(6): 1569-1581. |
| [5] | MENG Xiang-Yu, DIAO Deng-Chao, LIU Ya-Rui, LI Yun-Li, SUN Yu-Chen, WU Wei, ZHAO Wen, WANG Yu, WU Jian-Hui, LI Chun-Lian, ZENG Qing-Dong, HAN De-Jun, ZHENG Wei-Jun. Genetic analysis of high yield and yield stability characteristics of new wheat variety Xinong 877 [J]. Acta Agronomica Sinica, 2025, 51(5): 1261-1276. |
| [6] | XU Jie, XIA Lu-Lu, TANG Zhen-San, LI Wen-Li, ZHAO Tian-Tian, CHENG Li-Xiang, ZHANG Feng. Odor quality analysis of potato tuber after steaming and baking [J]. Acta Agronomica Sinica, 2025, 51(5): 1409-1420. |
| [7] | SONG Song-Quan, TANG Cui-Fang, CHENG Hong-Yan, WANG Cheng-Liang, YUAN Liang-Bing, ZUO Sheng. Endosperm development of cereal crops and its role in seed dormancy and germination [J]. Acta Agronomica Sinica, 2025, 51(5): 1133-1155. |
| [8] | SHEN Wen-Jie, CHEN Qing-Qing, HU Yi-Qun, ZHANG Ai-Fang, ZHANG Man-Yu. Study on the sensitivity of various physiological races of Magnaporthe oryzae to isoprothiolane in Anhui province, China [J]. Acta Agronomica Sinica, 2025, 51(5): 1338-1346. |
| [9] | MENG Fan-Qi, FANG Meng-Ying, LUO Yi, LU Lin, DONG Xue-Rui, WANG Ya-Fei, GUO Li-Na, YAN Peng, DONG Zhi-Qiang, ZHANG Feng-Lu. Effect of ethephon betaine salicylic acid mixture on heat resistance and yield of summer maize [J]. Acta Agronomica Sinica, 2025, 51(5): 1299-1311. |
| [10] | ZHAO Xi-Juan, ZHANG Fan, LIU Sheng-Xuan, QIN Jun, CHEN Hui-Lan, LIN Yuan, LUO Hong-Bing, LIU Yi, SONG Bo-Tao, HU Xin-Xi, WANG En-Shuang. Optimization of extraction methods for four endogenous hormones in potatoes and analysis of their content during the process of releasing dormancy in tubers [J]. Acta Agronomica Sinica, 2025, 51(4): 1050-1060. |
| [11] | ZHOU En-Qiang, MIAO Ya-Mei, ZHOU Yao, YAO Meng-Nan, ZHAO Na, WANG Yong-Qiang, ZHU Yu-Xiang, XUE Dong, LI Zong-Di, SHI Yu-Xin, LI Bo, WANG Kai-Hua, GU Chun-Yan, WANG Xue-Jun, WEI Li-Bin. Analysis of bZIP gene family and identification of seed development candidate genes in pea based on seed development transcriptome [J]. Acta Agronomica Sinica, 2025, 51(4): 914-931. |
| [12] | ZHANG Xiao-Li, LIU Xiao-Yan, XIA Wen-Wen, LI Jin. Functional analysis of the plasma membrane intrinsic protein gene SiPIP1;3 from Saussurea involucrata in tomato [J]. Acta Agronomica Sinica, 2025, 51(4): 863-872. |
| [13] | LI Hui-Min, XING Zhi-Peng, ZHANG Hai-Peng, WEI Hai-Yan, ZHANG Hong-Cheng, LI Guang-Yan. Application of chemical regulators and other cultivation measures in lodging resistance and high-yield cultivation of wheat [J]. Acta Agronomica Sinica, 2025, 51(4): 847-862. |
| [14] | JIN Gao-Rui, WU Xiao-Li, DENG Li, CHEN Yu-Ning, YU Bo-Lun, GUO Jian-Bin, DING Ying-Bin, LIU Nian, LUO Huai-Yong, CHEN Wei-Gang, HUANG Li, ZHOU Xiao-Jing, HUAI Dong-Xin, TAN Jia-Zhuang, JIANG Hui-Fang, REN Li, LEI Yong, LIAO Bo-Shou. Development and characterization of novel peanut genetic stocks with high oleic acid and enhanced resistance both to Aspergillus flavus infection and aflatoxin production [J]. Acta Agronomica Sinica, 2025, 51(3): 687-895. |
| [15] | JIN Xin-Xin, SONG Ya-Hui, SU Qiao, YANG Yong-Qing, LI Yu-Rong, WANG Jin. Identification and comprehensive evaluation of drought resistance in high oleic acid Jihua peanut varieties [J]. Acta Agronomica Sinica, 2025, 51(3): 797-811. |
|