欢迎访问作物学报,今天是
作物学报 ›› 2024, Vol. 50 ›› Issue (11): 2731-2741.doi: 10.3724/SP.J.1006.2024.44024
付佳祺1,2( ), 李世宽1,2, 谭萌慧1,2, 罗方3, 张传玲3, 刘祾悦3, 卢倩1,*(), 谷勇哲1,2,*()
), 李世宽1,2, 谭萌慧1,2, 罗方3, 张传玲3, 刘祾悦3, 卢倩1,*(), 谷勇哲1,2,*()
FU Jia-Qi1,2(), LI Shi-Kuan1,2, TAN Meng-Hui1,2, LUO Fang3, ZHANG Chuan-Ling3, LIU Ling-Yue3, LU Qian1,*(), GU Yong-Zhe1,2,*()
摘要:
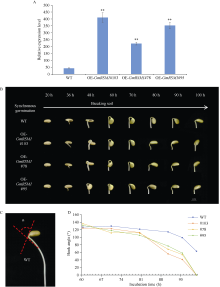
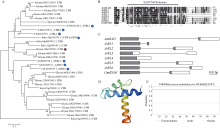
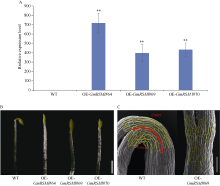
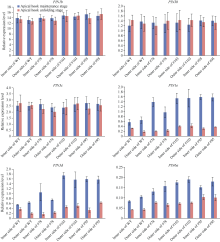
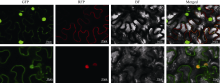
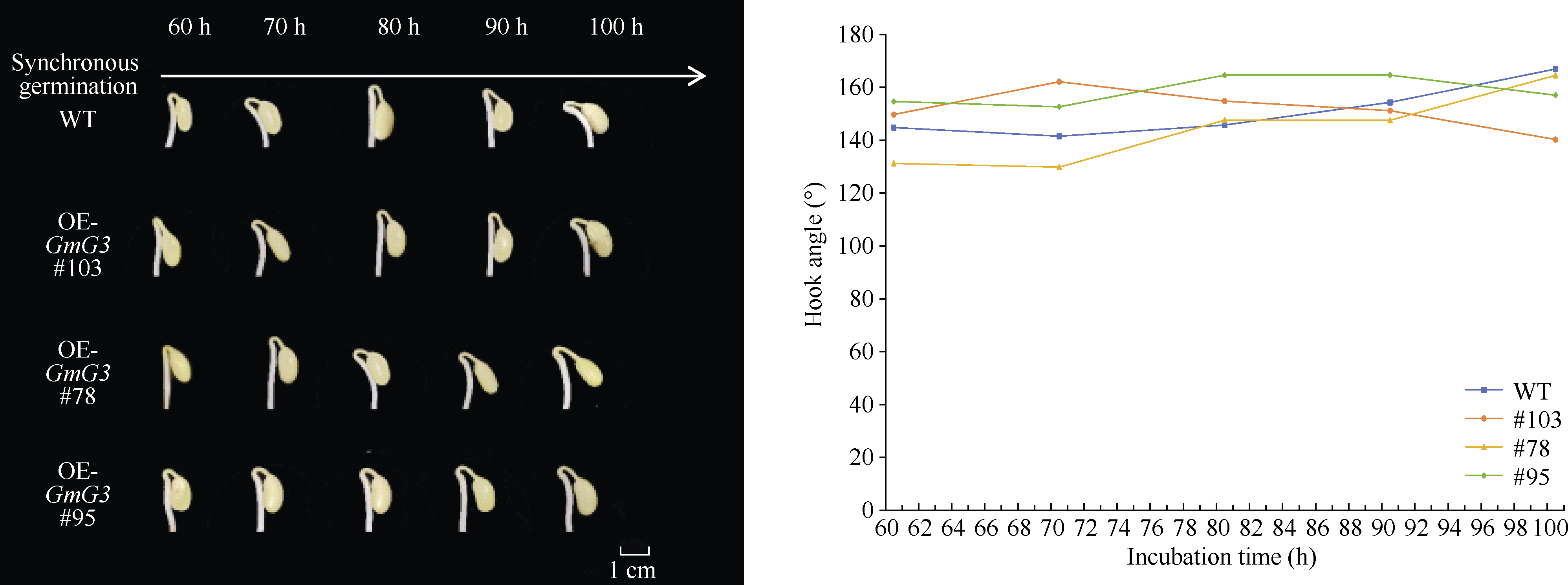
顶端弯钩是双子叶植物黄化幼苗短暂出现的独有结构, 尽管当前拟南芥顶端弯钩的模型网络已经相对透彻, 但在大豆中的研究却非常少。本试验构建pBAR-GmRSM1过表达载体, 通过遗传转化和阳性植株筛选, 得到3个拟南芥过表达纯合株系(OE-GmRSM1#64、#69和#70)和3个大豆纯合株系(OE-GmRSM1#103、#78和#95), 转化体的表达量均显著高于野生型。暗培养拟南芥和大豆, 过表达转化体的顶端弯钩消失或消失速度快于野生型。在扫描电镜下观察拟南芥野生型和转化体植株顶端弯钩部位细胞长度发现, 野生型在弯钩维持阶段弯钩内侧细胞长度小于外侧, 而转化体植株两侧细胞长度相同。分离大豆顶端弯钩内外两侧, 并通过检测PIN基因在大豆野生型与3个过表达转化体顶端弯钩的表达情况, 发现PIN1e、PIN3d和PIN6a三个基因在子叶出土时的顶端弯钩部位表达量显著高于顶端弯钩展开后的表达量, 而转化体中弯钩处这3个基因在弯钩维持阶段的表达量显著高于野生型, 但在展开后的表达量并无显著性差异。经过烟草瞬时表达, 确定GmRSM1蛋白定位在细胞核和细胞膜。因此, 顶端弯钩出现的原因是弯钩内外两侧细胞差异伸长, 因而导致下胚轴弯曲。GmRSM1可以通过正向调控生长素运输载体PIN1e、PIN3d和PIN6a缩短弯钩维持和展开时间, 验证了该基因对生长素转运的调控作用。本试验通过验证GmRSM1基因在顶端弯钩消失表型中的作用, 进一步完善了顶端弯钩的基因通路, 为后续研究奠定基础。
| [1] |
Veloccia A, Fattorini L, Della Rovere F, Sofo A, D’ Angeli S, Betti C, Falasca G, Altamura M M. Ethylene and auxin interaction in the control of adventitious rooting in Arabidopsis thaliana. J Exp Bot 2016, 67: 6445-6458.
doi: 10.1093/jxb/erw415 pmid: 27831474 |
| [2] | Shen X, Li Y, Pan Y, Zhong S. Activation of HLS1 by mechanical stress via ethylene-stabilized EIN3 is crucial for seedling soil emergence. Front Plant Sci, 2016, 7: 1571. |
| [3] | Peng Y, Zhang D, Qiu Y, Xiao Z, Ji Y, Li W, Xia Y, Wang Y, Guo H. Growth asymmetry precedes differential auxin response during apical hook initiation in Arabidopsis. J Integr Plant Biol 2022, 64: 5-22. |
| [4] |
Shi H, Liu R, Xue C, Shen X, Wei N, Deng X W, Zhong S. Seedlings transduce the depth and mechanical pressure of covering soil using COP1 and ethylene to regulate EBF1/EBF2 for soil emergence. Curr Biol, 2016, 26: 139-149.
doi: S0960-9822(15)01497-9 pmid: 26748855 |
| [5] | Silk W, Erickson R. Kinematics of hypocotyl curvature. Am J Bot, 1978, 65: 310-319. |
| [6] |
Weijers D, Nemhauser J, Yang Z. Auxin: small molecule, big impact. J Exp Bot, 2018, 69: 133-136.
doi: 10.1093/jxb/erx463 pmid: 29309681 |
| [7] | Schwark A, Schierle J. Interaction of ethylene and auxin in the regulation of hook growth: I. The role for ethylene in different growing regions of the hypocotyl hook of phaseolus vulgaris. J. Plant Physiol, 1992, 140: 562-570. |
| [8] | Žádníková P, Wabnik K, Abuzeineh A, Gallemi M, Van Der Straeten D, Smith R S, Inzé D, Friml J, Prusinkiewicz P, Benková E. A model of differential growth-guided apical hook formation in plants. Plant Cell, 2016, 28: 2464-2477. |
| [9] | Hamaguchi A, Yamashino T, Koizumi N, Kiba T, Kojima M, Sakakibara H, Mizuno T. A small subfamily of Arabidopsis RADIALIS-LIKE SANT/MYB genes: a link to HOOKLESS1- mediated signal transduction during early morphogenesis. Biosci Biotechnol Biochem, 2008, 72: 2687-2696. |
| [10] | Béziat C, Kleine-Vehn J. The road to auxin-dependent growth repression and promotion in apical hooks. Curr Biol, 2018, 28: R519-R525. |
| [11] |
Krecek P, Skupa P, Libus J, Naramoto S, Tejos R, Friml J, Zazímalová E. The PIN-FORMED (PIN) protein family of auxin transporters. Genome Biol, 2009, 10: 249.
doi: 10.1186/gb-2009-10-12-249 pmid: 20053306 |
| [12] |
Zádníková P, Petrásek J, Marhavy P, Raz V, Vandenbussche F, Ding Z, Schwarzerová K, Morita M T, Tasaka M, Hejátko J, Van Der Straeten D, Friml J, Benková E. Role of PIN-mediated auxin efflux in apical hook development of Arabidopsis thaliana. Development 2010, 137: 607-617.
doi: 10.1242/dev.041277 pmid: 20110326 |
| [13] |
Wang Y, Chai C, Valliyodan B, Maupin C, Annen B, Nguyen H T. Genome-wide analysis and expression profiling of the PIN auxin transporter gene family in soybean (Glycine max). BMC Genomics, 2015, 16: 951.
doi: 10.1186/s12864-015-2149-1 pmid: 26572792 |
| [14] |
Clough S J, Bent A F. Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J, 1998, 16: 735-743.
doi: 10.1046/j.1365-313x.1998.00343.x pmid: 10069079 |
| [15] | Wang Y, Li Z, Chen X, Gu Y, Zhang L, Qiu L. An efficient soybean transformation protocol for use with elite lines. Plant Cell Tissue Organ Cult, 2022, 151: 457-466. |
| [16] | Cheng G, Dong M, Xu Q, Peng L, Yang Z, Wei T, Xu J. Dissecting the molecular mechanism of the subcellular localization and cell-to-cell movement of the Sugarcane mosaic virus P3N-PIPO. Sci Rep, 2017, 7: 9868. |
| [17] | Wu Q, Li Y, Lyu M, Luo Y, Shi H, Zhong S. Touch-induced seedling morphological changes are determined by ethylene-regulated pectin degradation. Sci Adv, 2020, 6: eabc9294. |
| [18] |
Wang Y, Guo H. On hormonal regulation of the dynamic apical hook development. New Phytol, 2019, 222: 1230-1234.
doi: 10.1111/nph.15626 pmid: 30537131 |
| [19] | Li H, Johnson P, Stepanova A, Alonso J M, Ecker J R. Convergence of signaling pathways in the control of differential cell growth in Arabidopsis. Dev Cell 2004, 7: 193-204. |
| [20] |
Chai C, Wang Y, Valliyodan B, Nguyen H T. Comprehensive analysis of the soybean (Glycine max) GmLAX auxin transporter gene family. Front Plant Sci, 2016, 7: 282.
doi: 10.3389/fpls.2016.00282 pmid: 27014306 |
| [21] |
Baxter C E, Costa M M, Coen E S. Diversification and co-option of RAD-like genes in the evolution of floral asymmetry. Plant J, 2007, 52: 105-113.
pmid: 17672842 |
| [22] |
Costa M M, Fox S, Hanna A I, Baxter C, Coen E. Evolution of regulatory interactions controlling floral asymmetry. Development, 2005, 132: 5093-5101.
pmid: 16236768 |
| [23] |
Corley S B, Carpenter R, Copsey L, Coen E. Floral asymmetry involves an interplay between TCP and MYB transcription factors in Antirrhinum. Proc Natl Acad Sci USA, 2005, 102: 5068-5073.
pmid: 15790677 |
| [1] | 聂波涛, 刘德泉, 陈健, 崔正果, 侯云龙, 陈亮, 邱红梅, 王跃强. 北方春大豆品种农艺和品质性状分析与综合评价[J]. 作物学报, 2024, 50(9): 2248-2266. |
| [2] | 孙现军, 胡正, 姜雪敏, 王世佳, 陈向前, 张惠媛, 张辉, 姜奇彦. 大豆种质资源苗期耐盐性鉴定评价与筛选[J]. 作物学报, 2024, 50(9): 2179-2186. |
| [3] | 刘欣玥, 郭潇阳, 王欣茹, 辛大伟, 关荣霞, 邱丽娟. 大豆萌发期耐盐性鉴定方法建立及耐盐大豆资源筛选[J]. 作物学报, 2024, 50(8): 2122-2130. |
| [4] | 李晓菲, 高华伟, 广慧, 石宇欣, 谷勇哲, 齐照明, 邱丽娟. 大豆种质资源萌发期耐莠去津鉴定评价及优异种质筛选[J]. 作物学报, 2024, 50(7): 1699-1709. |
| [5] | 张红梅, 张威, 王琼, 贾倩茹, 孟珊, 熊雅文, 刘晓庆, 陈新, 陈华涛. 大豆籽粒Ve含量的全基因组关联分析[J]. 作物学报, 2024, 50(5): 1223-1235. |
| [6] | 苗龙, 舒阔, 李娟, 黄茹, 王业杏, Soltani Muhammad YOUSOF, 许竞好, 吴传磊, 李佳佳, 王晓波, 邱丽娟. 大豆根茎过渡区弯曲突变体Mrstz的鉴定与基因定位[J]. 作物学报, 2024, 50(5): 1091-1103. |
| [7] | 王亚琪, 徐海风, 李曙光, 傅蒙蒙, 余希文, 赵志鑫, 杨加银, 赵团结. 大豆类病变皱叶突变体NT301遗传分析和2对基因定位[J]. 作物学报, 2024, 50(4): 808-819. |
| [8] | 王琼, 朱宇翔, 周密密, 张威, 张红梅, 陈新, 陈华涛, 崔晓艳. 大豆叶型性状全基因组关联分析与候选基因鉴定[J]. 作物学报, 2024, 50(3): 623-632. |
| [9] | 刘薇, 王玉斌, 李伟, 张礼凤, 徐冉, 王彩洁, 张彦威. 过量表达大豆异丙基苹果酸脱氢酶基因GmIPMDH促进植株开花和生长[J]. 作物学报, 2024, 50(3): 613-622. |
| [10] | 宋健, 熊亚俊, 陈伊洁, 徐瑞新, 刘康林, 郭庆元, 洪慧龙, 高华伟, 谷勇哲, 张丽娟, 郭勇, 阎哲, 刘章雄, 关荣霞, 李英慧, 王晓波, 郭兵福, 孙如建, 闫龙, 王好让, 姬月梅, 常汝镇, 王俊, 邱丽娟. 大豆巢式关联作图(NAM)群体构建及花色和种皮色遗传分析[J]. 作物学报, 2024, 50(3): 556-575. |
| [11] | 李世宽, 洪慧龙, 付佳祺, 谷勇哲, 孙如建, 邱丽娟. BSA-Seq结合RNA-Seq技术挖掘大豆叶片提前黄化衰老基因[J]. 作物学报, 2024, 50(2): 294-309. |
| [12] | 谭丹, 陈家婷, 郜钰, 张晓军, 李欣, 闫贵云, 李锐, 陈芳, 常利芳, 张树伟, 郭慧娟, 畅志坚, 乔麟轶. 小麦穗型相关生长素通路基因发掘及TaARF23-A与小穗数关联分析[J]. 作物学报, 2024, 50(2): 506-513. |
| [13] | 方然, 袁丽媚, 王玉林, 芦思佳, 孔凡江, 刘宝辉, 孔令平. 生育期基因E1~E4不同突变组合对大豆纬度适应性的影响[J]. 作物学报, 2024, 50(12): 3013-3024. |
| [14] | 王子然, 鲁一薇, 杨婧怡, 王成龙, 宋亚萍, 马金虎. 外源水杨酸对镉胁迫下大豆生理特性和抗逆基因表达的影响[J]. 作物学报, 2024, 50(11): 2883-2895. |
| [15] | 李宛鸿, 胡冰霜, 孙晓丽, 才晓溪, 孙明哲. 过表达野生大豆耐盐碱基因GsGSTU13提高了水稻苗期耐盐碱性[J]. 作物学报, 2024, 50(10): 2458-2467. |
|
||